
RESEARCH
フォトトロピンを介した光応答
植物は光合成を通じて太陽からの光エネルギーを化学エネルギーに変換し、地球上の生命活動の根幹を支えています。さらに植物は、光を光合成に必要なエネルギー源として利用するのみならず、「環境シグナル」としても利用し、自身の成長を最適化しています。
フォトトロピンは植物に特有な青色光受容体キナーゼであり、光屈性、葉緑体光定位運動、気孔開口、葉の展開など、光合成機能の最適化に関わる多様な光応答を制御しています。実際に私たちは、植物がフォトトロピンを介して青色光をシグナルとして利用することで、成長を約3倍促進することを実証しました(Plant Cell 2005)。しかし、そのシグナル伝達の仕組みについては、フォトトロピンによりリン酸化される基質タンパク質の実体をはじめとして、いまだ多くの未解明な点が残されています。

図1 フォトトロピンの構造と光応答。フォトトロピンはN末端側に光受容に関わる2つのLOVドメインを、C末端側にSer/Thrキナーゼドメインをもつ光受容体キナーゼである。青色光の受容によって活性化したフォトトロピンは、細胞内シグナル伝達を介して、多様な光応答を誘導する。これらの諸反応は、植物の光受容や二酸化炭素吸収を高め、光合成・成長を促進する。
青色光に応答した気孔開口の分子メカニズム
私たちは、植物における環境応答・シグナル伝達のモデル系として、青色光に応答した気孔開口のシグナル伝達機構の解明に取り組んでいます。これまでに、赤外線サーモグラフィを用いて気孔開口を視覚的に検出する系を構築し、モデル植物シロイヌナズナを対象とした大規模変異体スクリーニングにより、気孔開口の必須因子であるBLUS1キナーゼを同定しました(Nature Commun 2013)。さらに孔辺細胞プロトプラストを用いたリン酸化プロテオーム解析により、BLUS1がフォトトロピンによって直接リン酸化されることを示し、長らく不明であったフォトトロピンのリン酸化基質を初めて同定しました。加えて、BLUS1の活性制御機構の解明(Plant Cell 2021)や、BLUS1下流のシグナル伝達にprotein phosphatase1 (PP1)が関与することの解明(PNAS 2006)を進めるとともに、細胞膜H+-ATPaseの光活性化にはC末端自己阻害領域に存在する2カ所のThr残基のリン酸化が必須であることを明らかにしました(Nature Commun 2024)。
さらに私たちは、フォトトロピンの第2のリン酸化基質としてCBCキナーゼを同定し、CBCが気孔閉鎖の鍵因子であるアニオンチャネルを不活性化することで、気孔開口を促進することを明らかにしました(Nature Commun 2017)。CBCはフォトトロピンのみならず、CO2センサーであるHT1によってもリン酸化され、単一分子内で青色光シグナルとCO2シグナル伝達を統合するクロストークポイントとして機能することを示しました。さらに近年、私たちはフォトトロピンの第3のリン酸化基質としてWDR48タンパク質を同定し、本因子が孔辺細胞葉緑体におけるデンプン分解を介して気孔開口を促進することを明らかにしました(Nature Commun 2026)。
これらの研究により、フォトトロピンは孔辺細胞内で多様な因子を同時に制御し精巧なシグナル伝達ネットワークを形成することで、応答を厳密且つ統合的に制御する新たなシグナル伝達モデルが見えてきました。
現在、生化学、分子遺伝学、分子生物学、構造生物学、イメージング解析、人工知能・深層学習など、多角的なアプローチを融合し、シグナル伝達に関わる新たな重要因子・メカニズムの解明を進めています。フォトトロピンが制御する「光シグナル伝達ネットワーク」の全貌を明らかにすることで、植物がもつ巧妙な環境応答・シグナル伝達の仕組みの解明を目指しています。
さらに、乾燥ストレスに応答して合成されるアブシシン酸(ABA)や、様々な生物的・非生物的ストレス条件下で生成される活性酸素種(PNAS 2019)、さらには概日時計など、多様な外的・内的要因と青色光シグナル伝達とのクロストークについても研究を展開しています。これにより、植物が変動する環境下で気孔開度を最適化させるメカニズムの解明を目指しています。
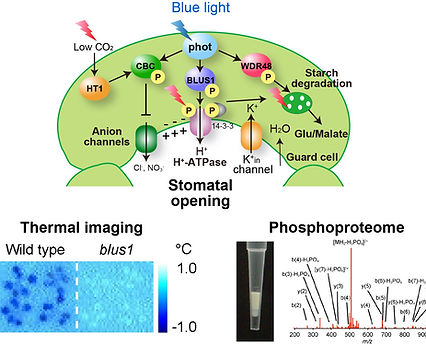
図2 (上段)青色光による気孔開口のシグナル伝達ネットワークモデル。(下段左)赤外線サーモグラフィを用いて青色光による気孔開口を視覚的に検出した熱画像。野生株では青色光に応じて気孔が開口するため葉温が低下するが、blus1変異体では気孔が開口せず葉温は低下しない。(下段右)リン酸化プロテオームでは細胞内のタンパク質のリン酸化修飾を網羅的に解析することができる。赤外線サーモグラフィを用いた変異体スクリーニングとリン酸化プロテオーム解析は、シグナル伝達ネットワーク研究の強力なツールとなっている。
Selected Publications:
-
Yamauchi S, Fuji S, Ikuta H, Sugiyama N, Kodama Y, Distefano L, Fujii H, Yamashita K, Takase H, Nomoto M, Tada Y, Umezawa T, Hotta K, Santelia D, Shimazaki K, Takemiya A (2026) Phosphorylation of WDR48 by phototropins drives starch degradation to promote stomatal opening. Nature Commun.
-
Fuji S, Yamauchi S, Sugiyama N, Kohchi T, Nishihama R, Shimazaki K, Takemiya A (2024) Light-induced stomatal opening requires phosphorylation of the autoinhibitory domain of plasma membrane H+-ATPase. Nature Commun 15: 1195.
-
Hosotani S, Yamauchi S, Kobayashi H, Fuji S, Koya S, Shimazaki K, Takemiya A (2021) A BLUS1 kinase signal and a decrease in intercellular CO2 concentration are necessary for stomatal opening in response to blue light. Plant Cell 33: 1813-1827.
-
Yamauchi S, Mano S, Oikawa K, Hikino K, Teshima KM, Kimori Y, Nishimura M, Shimazaki K, Takemiya A (2019) Autophagy controls reactive oxygen species homeostasis in guard cells that is essential for stomatal opening. Proc Natl Acad Sci USA 116: 19187-19192.
-
Hiyama A, Takemiya A, Munemasa S, Okuma E, Sugiyama N, Tada Y, Murata Y, Shimazaki K (2017) Blue light and CO2 signals converge to regulate light-induced stomatal opening. Nature Commun 8: 1284.
-
Takemiya A, Sugiyama N, Fujimoto H, Tsutsumi T, Yamauchi S, Hiyama A, Tada Y, Christie JM, Shimazaki K (2013) Phosphorylation of BLUS1 kinase by phototropins is a primary step in stomatal opening. Nature Commun 4: 2094.
-
Takemiya A, Kinoshita T, Asanuma M, Shimazaki K (2006) Protein phosphatase 1 positively regulates stomatal opening in response to blue light in Vicia faba. Proc Natl Acad Sci USA 103: 13549-13554.
-
Takemiya A, Inoue S, Doi M, Kinoshita T, Shimazaki K (2005) Phototropins promote plant growth in response to blue light in low light environments. Plant Cell 17: 1120-1127.